À propos de ce travail
Ce projet a été réalisé dans le cadre du cours Adaptation et Phylogénie du master BEE, parcours SEP, à Sorbonne Université et au Muséum national d’Histoire naturelle.
Il propose une analyse phylogénétique et morpho-fonctionnelle de l’ornithorynque (Ornithorhynchus anatinus) afin d’interroger l’origine de ses adaptations semi-aquatiques : relèvent-elles d’une niche ancienne conservée au sein des monotrèmes, ou d’une spécialisation secondaire convergente ?
Contexte biologique
Les monotrèmes constituent l’ordre des mammifères ovipares. Ils sont aujourd’hui répartis entre l’Australie, la Tasmanie et la Nouvelle-Guinée.
Ils présentent une combinaison singulière de caractères plésiomorphes, conservés ou dérivés par rapport aux autres mammifères, parmi lesquels :
- la présence d’un cloaque ;
- l’absence de mamelons, les jeunes se nourrissant du lait sécrété par des glandes mammaires situées sur la peau ;
- une ceinture scapulaire très développée ;
- une posture semi-étalée ;
- la présence d’un éperon tarsien chez le mâle.
Les monotrèmes constituent ainsi un groupe-clef pour comprendre l’évolution des mammifères. Ils sont parfois qualifiés de « fossiles vivants », une expression pratique, mais réductrice, car leur morphologie combine des caractères hérités anciens et des spécialisations propres.
Aujourd’hui, les monotrèmes sont représentés par cinq espèces réparties entre deux grands profils écologiques : les échidnés, terrestres et fouisseurs, et l’ornithorynque, semi-aquatique.

Crédit photo : Joshua Earle, Unsplash
Données et méthodes
Taxons étudiés
-
Monotrèmes :
- Tachyglossus aculeatus (échidné à nez court) ;
- Zaglossus bruijni (échidné à long nez) ;
- Ornithorhynchus anatinus (ornithorynque) ;
-
Marsupiaux :
- Macropus rufus (kangourou roux) ;
- Didelphis marsupialis (oppossum commun) ;
-
Placentaires :
- Homo sapiens (être humain) ;
- Felis catus (chat domestique) ;
- Loxodonta africana (éléphant de savane d’Afrique) ;
- Ovis aries (mouton) ;
- Equus caballus (cheval) ;
- Choloepus hoffmanni (paresseux) ;
- Myotis lucifugus (petite chauve-souris brune) ;
- Orycteropus afer (oryctérope) ;
- Ondatra zibethicus* (rat musqué) ;
- Castor canadensis* (castor du Canada) ;
- Lutra lutra* (loutre d’Europe) ;
- Zalophus californianus* (otarie de Californie) ;
-
Groupe externe :
- Anas platyrhynchos (canard colvert).
* taxons semi-aquatiques
Taxons fossiles
-
Monotrèmes et apparentés :
- Obdurodon dicksoni ;
- Teinolophos trusleri ;
- Steropodon galmani ;
- Kollikodon ritchiei ;
- Monotrematum sudamericanum ;
-
Mammaliformes :
- Castorocauda lutrasimilis* ;
- Morganucodon watsoni.
* taxon semi-aquatique
Marqueurs moléculaires
Pour ce travail, les marqueurs suivants ont été étudiés :
- cytochrome B (
CYTB) ; - cytochrome c oxydase I (
COX1) ; - ARN ribosomique 16S (
16S) ; - gène activateur de la recombinaison V(D)J, codant la protéine RAG1 (
RAG1).
Les marqueurs ont été alignés, puis concaténés pour les analyses.
Caractères morpho-fonctionnels
Une matrice de caractères morpho-fonctionnels a été construite afin de comparer les taxons selon deux ensembles de traits : des caractères mammaliens généraux, incluant des caractères reproducteurs propres aux grands clades de mammifères, et des caractères associés à la spécialisation semi-aquatique.
Caractères mammaliens généraux
- poils ;
- glandes mammaires ;
- oreille moyenne à trois osselets ;
- diphyodontie ;
- hétérodontie ;
- oviparité ;
- cloaque ;
- mamelons ;
- marsupium ;
- placenta ;
- dents adultes ;
- bec kératinisé ;
- éperon tarsien ;
- venin ;
- température corporelle < 34°C ;
- rostre spécialisé ;
- fouissage ;
- queue aplatie ;
- interclavicule ;
- posture semi-étalée ;
- os épipubiens.
Caractères associés à la spécialisation semi-aquatique
- membres palmés ;
- membres transformés en nageoires ;
- corps hydrodynamique ;
- mode de propulsion ;
- queue spécialisée ;
- fourrure dense ;
- réduction de l’oreille externe ;
- fermeture des yeux ;
- fermeture des oreilles ;
- recherche alimentaire sous l’eau ;
- électroréception aquatique ;
- densification humérale ;
- plongeon prolongé ;
- réduction de la locomotion terrestre ;
- ostéosclérose.
Analyses phylogénétiques et morpho-fonctionnelles
Les analyses ont été organisées autour de quatre étapes complémentaires.
Reconstruction d’un arbre phylogénétique de référence
Les séquences moléculaires disponibles pour les marqueurs CYTB, COX1, 16S et RAG1 ont été alignées, puis concaténées afin de reconstruire un arbre phylogénétique de référence.
Cet arbre permet de replacer l’ornithorynque dans le cadre évolutif général des mammifères étudiés et de disposer d’un support pour les analyses comparatives ultérieures.
L’arbre a été construit par inférence bayésienne après alignement et concaténation des marqueurs moléculaires, en utilisant un modèle GTR + I + Γ par partition. Deux runs indépendants de quatre chaînes ont été lancés, avec un burn-in de 25 %. La convergence a été contrôlée avec l’ASDSF (moyenne = 0,003865) et le PSRF (moyenne = 1,000). L’arbre consensus obtenu à partir de l’alignement concaténé a ensuite été utilisé comme arbre de référence.
Analyse morpho-fonctionnelle par PCoA
Les caractères morpho-fonctionnels ont été codés sous forme de matrice, puis projetés dans un espace multivarié à l’aide d’une PCoA.
Deux analyses ont été distinguées :
- une PCoA fondée sur les caractères mammaliens généraux ;
- une PCoA fondée sur les caractères associés à la spécialisation semi-aquatique.
Cette distinction permet de comparer deux signaux différents : d’une part, le signal phylogénétique lié aux grands clades de mammifères ; d’autre part, le signal fonctionnel lié au mode de vie semi-aquatique.
Reconstruction ancestrale du mode de vie
Une reconstruction ancestrale discrète du mode de vie a été réalisée sur l’arbre phylogénétique afin d’estimer l’état le plus probable au nœud des monotrèmes.
L’objectif était de déterminer si le mode de vie semi-aquatique de l’ornithorynque pouvait être interprété comme un état ancien conservé chez les monotrèmes, ou comme une spécialisation plus récente propre à la lignée de l’ornithorynque.
Analyse exploratoire des taxons fossiles
Enfin, plusieurs taxons fossiles ont été intégrés de manière exploratoire, notamment Obdurodon dicksoni, afin d’évaluer dans quelle mesure les fossiles modifient l’interprétation du signal semi-aquatique.
Cette étape est importante car les taxons actuels seuls opposent fortement les échidnés terrestres et l’ornithorynque semi-aquatique, ce qui rend l’état ancestral difficile à interpréter.
Résultats
Les résultats doivent être lus comme la comparaison de deux signaux : un signal phylogénétique, lié à la position des monotrèmes parmi les mammifères, et un signal morpho-fonctionnel, lié aux adaptations semi-aquatiques.
Arbre phylogénétique de référence
L’arbre phylogénétique obtenu après inférence bayésienne est présenté en Figure 1. Il fournit un cadre phylogénétique cohérent pour comparer les profils morpho-fonctionnels des taxons étudiés.
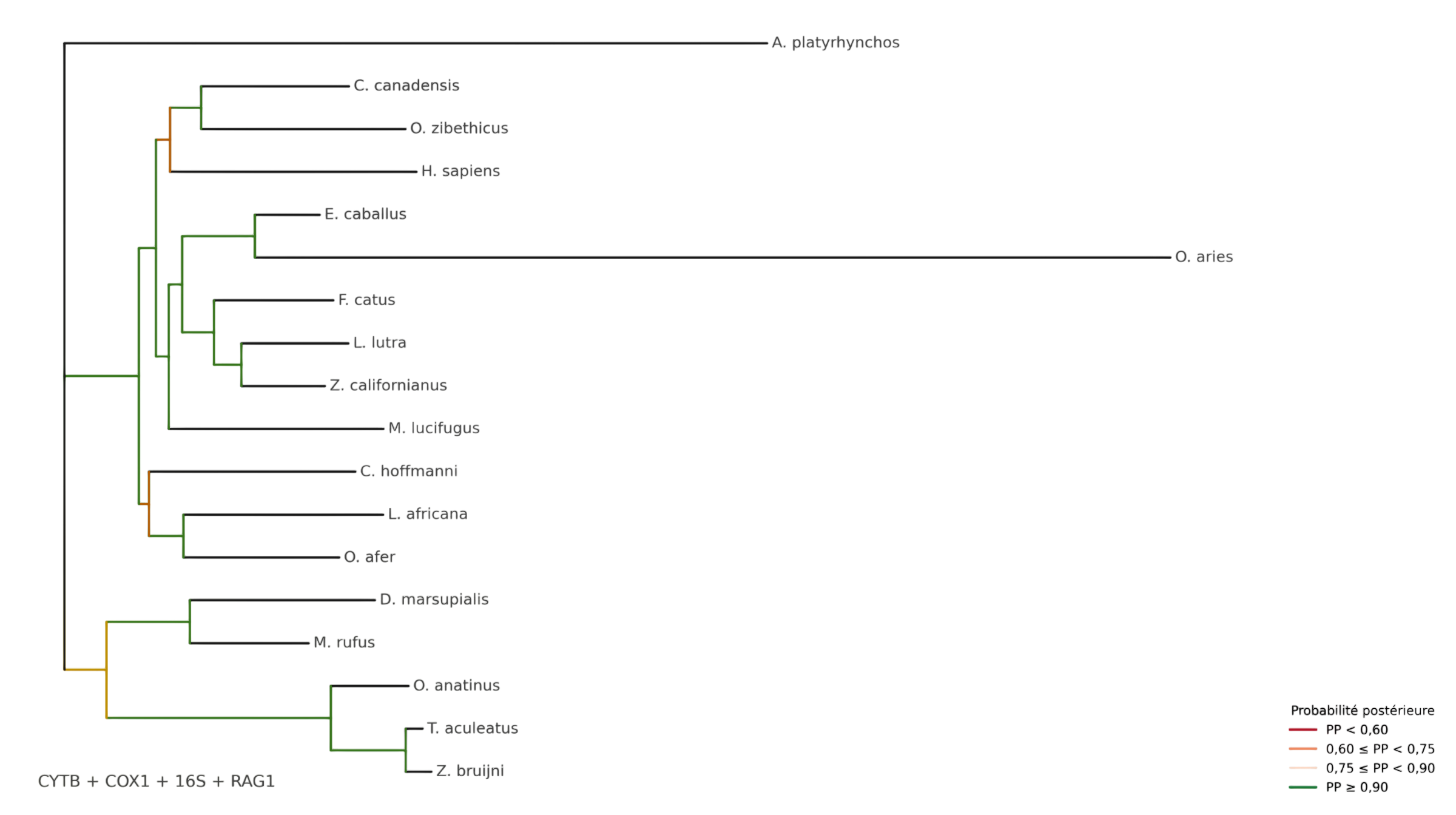
Figure 1 - Arbre phylogénétique de référence
Le clade des monotrèmes est bien récupéré et regroupe les trois taxons attendus (O. anatinus, T. aculeatus et Z. bruijni).
Cependant, les relations profondes entre les placentaires sont moins stables.
Signal mammalien général
La PCoA sur caractères mammaliens généraux montre une distance réduite entre les trois taxons monotrèmes. Elle est présentée en Figure 2.
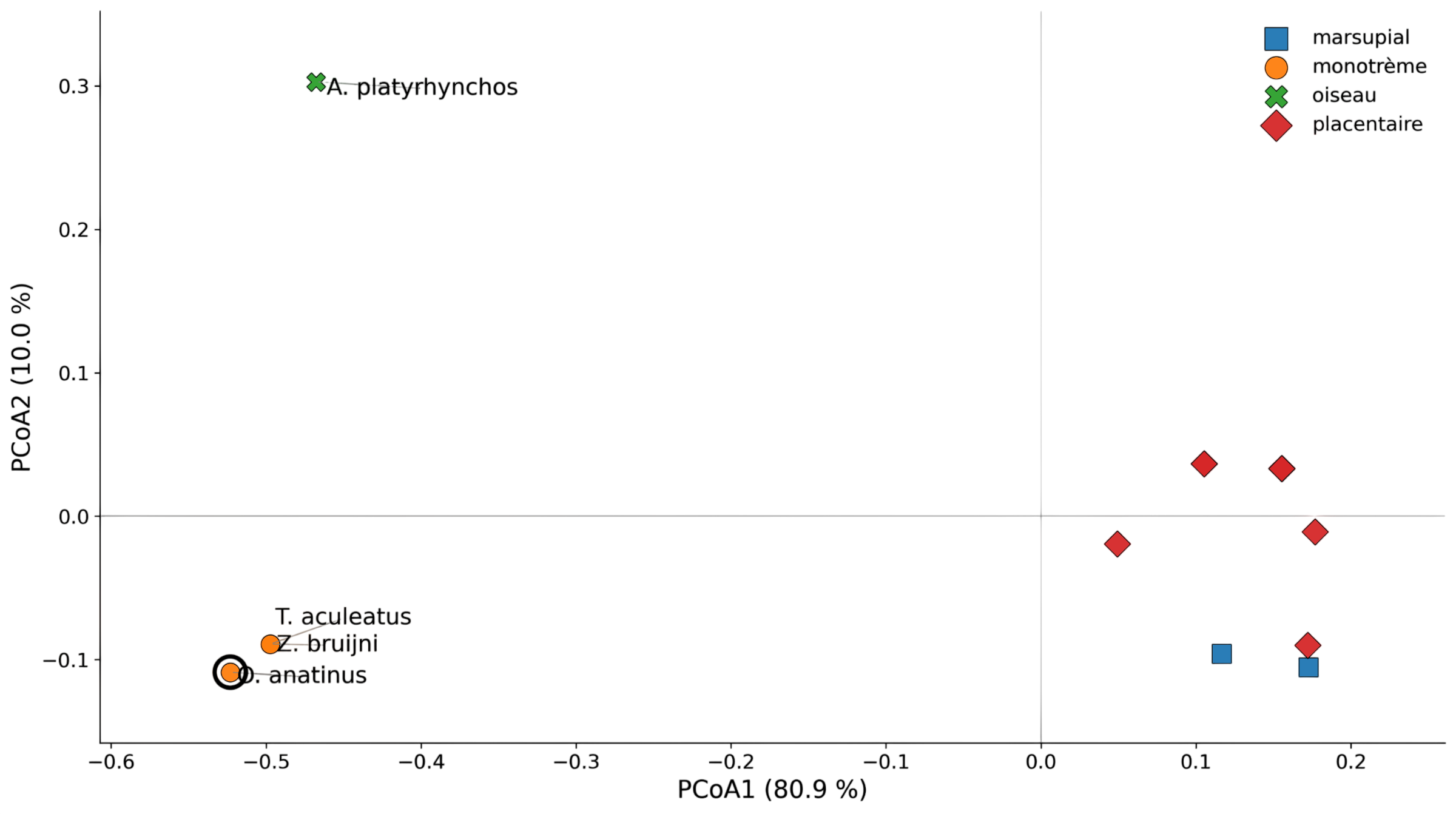
Figure 2 - PCoA sur les caractères mammaliens généraux
Cela indique que le clade des monotrèmes est bien supporté par les caractères mammaliens généraux.
Signal semi-aquatique
La Figure 3 présente les résultats de la PCoA sur les caractères morpho-fonctionnels semi-aquatiques.
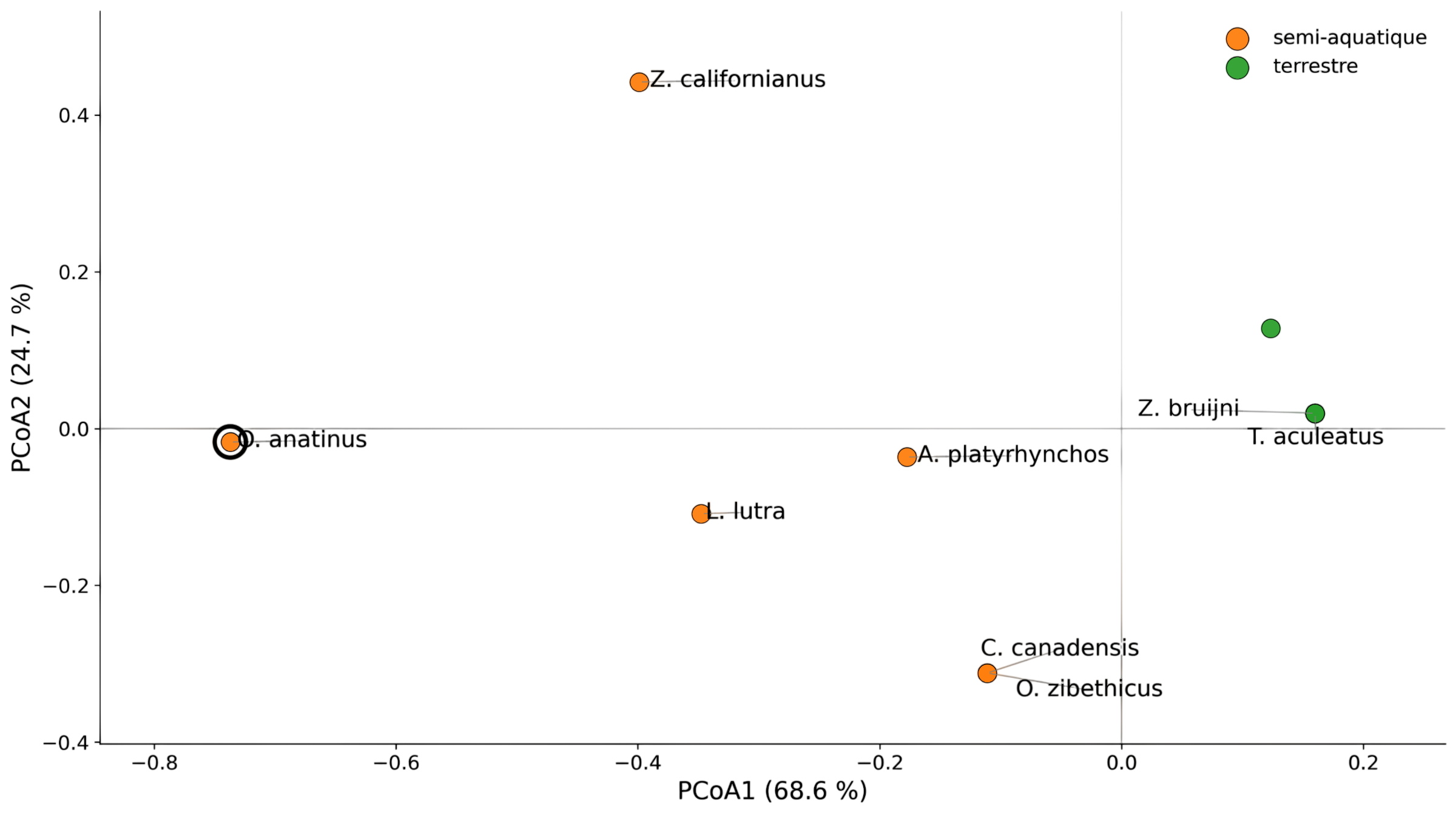
Figure 3 - PCoA sur les caractères morpho-fonctionnels semi-aquatiques
Les échidnés sont éloignés de l’ornithorynque dans l’espace morpho-fonctionnel semi-aquatique. Les monotrèmes actuels ne partagent donc pas tous le même profil écologique et fonctionnel.
Les mammifères semi-aquatiques placentaires ne forment pas non plus un bloc unique : Zalophus californianus, Lutra lutra, Castor canadensis et Ondatra zibethicus sont dispersés dans l’espace morpho-fonctionnel. Le terme « semi-aquatique » ne correspond donc pas à un syndrome morphologique unique.
L’ornithorynque forme un pôle propre, suggérant que son signal semi-aquatique ne correspond pas à une simple convergence avec les mammifères placentaires semi-aquatiques, mais à une combinaison spécifique de caractères.
Reconstruction ancestrale sans fossiles
La reconstruction ancestrale discrète du mode de vie, réalisée à partir des taxons actuels seuls, ne permet pas de trancher clairement entre un état terrestre et un état semi-aquatique/aquatique au nœud des monotrèmes.
Ce résultat reflète le contraste entre les monotrèmes actuels : les échidnés sont terrestres et fouisseurs, tandis que l’ornithorynque est semi-aquatique. Les taxons actuels seuls ne suffisent donc pas à distinguer clairement une niche ancienne conservée d’une spécialisation secondaire propre à la lignée de l’ornithorynque.
Apport d’Obdurodon dicksoni
L’ajout d’Obdurodon dicksoni déplace modérément l’estimation du nœud des monotrèmes vers l’état semi-aquatique/aquatique.
L’intégration d’Obdurodon dicksoni renforce donc l’hypothèse d’un signal semi-aquatique ancien au sein des Ornithorhynchidae, sans démontrer que cet état caractérisait l’ensemble des monotrèmes ancestraux.
Position morpho-fonctionnelle du nœud monotrème
La position morpho-fonctionnelle du nœud monotrème a été estimée dans l’espace PCoA semi-aquatique (Figure 4).
Pour cela, les coordonnées des taxons sur les axes PCoA1 et PCoA2 ont été utilisées comme deux traits continus. Leur valeur au nœud des monotrèmes a ensuite été estimée par maximum de vraisemblance, sous un modèle brownien d’évolution des traits continus.
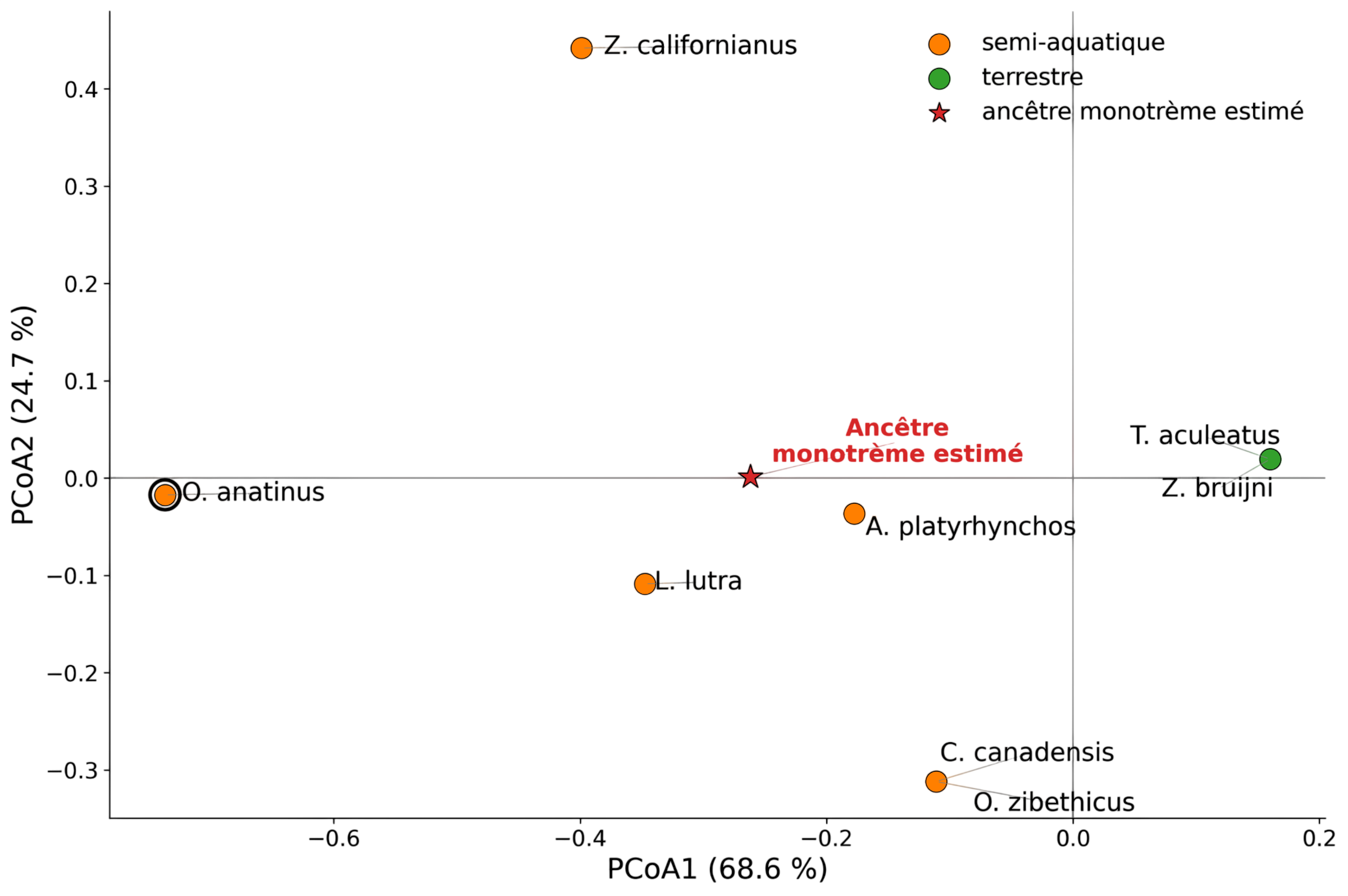
Figure 4 - Position morpho-fonctionnelle du nœud monotrème estimé dans l’espace PCoA semi-aquatique
Le nœud monotrème estimé occupe une position intermédiaire dans l’espace morpho-fonctionnel semi-aquatique. Il ne correspond pas au profil très spécialisé de l’ornithorynque actuel, mais ne se rapproche pas non plus strictement du profil terrestre des échidnés.
Cette position intermédiaire est compatible avec une mosaïque morpho-fonctionnelle : le signal reconstruit au nœud monotrème ne suggère ni un état pleinement comparable à celui de l’ornithorynque actuel, ni un état simplement assimilable aux échidnés terrestres.
Projection exploratoire des fossiles
La Figure 5 présente la PCoA exploratoire réalisée sur une matrice fossile réduite, limitée aux caractères pouvant être codés de manière suffisamment fiable chez les taxons fossiles.
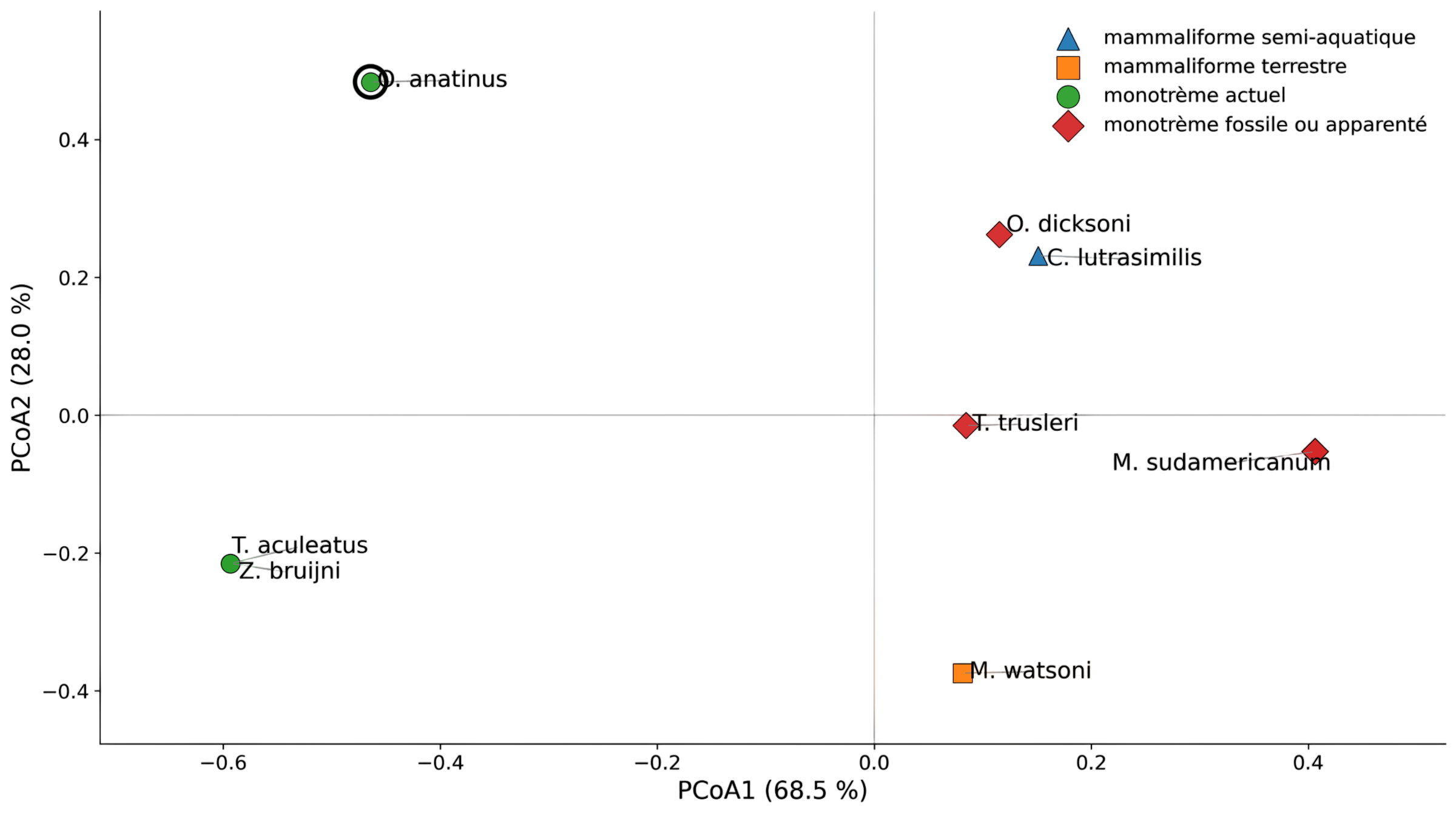
Figure 5 - PCoA fossile exploratoire
Dans cet espace morpho-fonctionnel, Obdurodon dicksoni se projette à proximité du pôle semi-aquatique, proche de Castorocauda lutrasimilis, un mammaliaforme semi-aquatique convergent. Cette position est compatible avec l’hypothèse d’une spécialisation semi-aquatique ancienne au sein de la lignée des Ornithorhynchidae.
À l’inverse, Morganucodon watsoni sert ici de point de comparaison terrestre. Sa position permet de mieux situer les fossiles dans l’espace morpho-fonctionnel, même si l’analyse reste qualitative.
Les autres monotrèmes fossiles inclus dans l’analyse, notamment Teinolophos trusleri, Monotrematum sudamericanum et les formes apparentées plus fragmentaires, sont surtout informatifs sur la diversité dentaire et phylogénétique des monotrèmes. Leur intérêt est plus limité pour inférer directement l’écologie, car les caractères liés au mode de vie semi-aquatique sont souvent absents ou difficilement codables.
Cette projection doit donc être interprétée avec prudence : elle suggère un signal cohérent avec une spécialisation ancienne de la lignée de l’ornithorynque, mais le codage fossile incomplet empêche d’en faire un argument conclusif à lui seul.
Tests complémentaires
Des tests complémentaires ont été réalisés afin de préciser la nature des signaux observés dans les matrices de caractères.
Le signal phylogénétique a d’abord été évalué avec le λ de Pagel. Pour les caractères mammaliens généraux, λ est proche de 1 et très significatif, ce qui indique que cette matrice suit fortement la structure phylogénétique : les espèces proches phylogénétiquement tendent à partager davantage de caractères. Ce résultat est cohérent avec la PCoA fondée sur les caractères mammaliens généraux, qui regroupe les monotrèmes entre eux.
À l’inverse, pour les caractères associés au mode de vie semi-aquatique, λ est proche de 0 et non significatif. Le signal morpho-fonctionnel semi-aquatique n’est donc pas principalement expliqué par la proximité phylogénétique.
Une PERMANOVA réalisée sur les distances de Gower a ensuite été utilisée pour tester la séparation entre taxons terrestres et semi-aquatiques. Le résultat n’est pas significatif, ce qui indique que le signal semi-aquatique ne se réduit pas non plus à deux groupes homogènes.
Ces tests confirment donc une double dissociation : les caractères mammaliens généraux conservent un signal phylogénétique fort, tandis que les caractères semi-aquatiques traduisent un signal morpho-fonctionnel plus complexe, non réductible ni à la phylogénie, ni à une opposition binaire simple entre formes terrestres et semi-aquatiques.
Conclusion
Ce projet met en évidence une dissociation entre deux niveaux de signal.
D’un côté, les caractères mammaliens généraux soutiennent clairement le regroupement des monotrèmes et conservent un signal phylogénétique fort. De l’autre, les caractères associés au mode de vie semi-aquatique isolent l’ornithorynque des échidnés actuels et ne se réduisent pas à une simple convergence avec les mammifères placentaires semi-aquatiques.
La reconstruction ancestrale sans fossiles ne permet pas de trancher entre conservation ancienne et spécialisation secondaire. L’ajout d’Obdurodon dicksoni déplace cependant l’interprétation vers un signal semi-aquatique plus ancien au sein des Ornithorhynchidae.
L’ornithorynque apparaît ainsi comme une mosaïque évolutive : profondément inscrit dans l’histoire des monotrèmes, mais porteur d’une combinaison morpho-fonctionnelle propre liée à son mode de vie semi-aquatique.
Limites et perspectives
Plusieurs limites doivent être prises en compte dans l’interprétation des résultats.
La première concerne l’échantillonnage. Les monotrèmes actuels sont peu nombreux, et les taxons fossiles disponibles restent fragmentaires. Cette faible densité taxonomique limite la capacité à distinguer clairement une conservation ancienne du mode de vie semi-aquatique d’une spécialisation secondaire propre à la lignée de l’ornithorynque.
La deuxième limite concerne le codage morpho-fonctionnel. Certains caractères peuvent être corrélés entre eux, notamment lorsqu’ils participent à un même complexe fonctionnel lié à la nage, à la plongée ou à la recherche alimentaire sous l’eau. Par ailleurs, plusieurs traits ne peuvent pas être codés chez les fossiles lorsqu’ils ne sont pas directement observables, en particulier pour les caractères liés aux tissus mous ou au comportement.
Le cadre phylogénétique constitue également une limite. L’arbre de référence a été construit à partir de données moléculaires disponibles pour les taxons actuels. Les taxons fossiles ne pouvant pas être intégrés automatiquement dans cette inférence moléculaire, Obdurodon dicksoni a été greffé manuellement dans l’arbre, sur la base de sa position phylogénétique attendue au sein des Ornithorhynchidae.
Une perspective importante serait de décomposer le signal semi-aquatique caractère par caractère, afin d’identifier quels traits contribuent le plus à la position particulière de l’ornithorynque et d’Obdurodon dicksoni. Une telle approche nécessiterait toutefois des fossiles plus complets, notamment pour les éléments post-crâniens, qui sont essentiels pour discuter les adaptations locomotrices et aquatiques.

Crédit photo : Wildlife Warriors
Références
Archer, M., Flannery, T. F., Ritchie, A., & Molnar, R. E. (1985). First Mesozoic mammal from Australia—An early Cretaceous monotreme. Nature, 318(6044), 363–366. https://doi.org/10.1038/318363a0
Flannery, T. F., Rich, T. H., Vickers-Rich, P., Ziegler, T., Veatch, E. G., & Helgen, K. M. (2022). A review of monotreme (Monotremata) evolution. Alcheringa: An Australasian Journal of Palaeontology, 46(1), 3–20. https://doi.org/10.1080/03115518.2022.2025900
Ji, Q., Luo, Z.-X., Yuan, C.-X., & Tabrum, A. R. (2006). A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science, 311(5764), 1123–1127. https://doi.org/10.1126/science.1123026
Musser, A. M., & Archer, M. (1998). New information about the skull and dentary of the Miocene platypus Obdurodon dicksoni, and a discussion of ornithorhynchid relationships. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 353(1372), 1063–1079. https://doi.org/10.1098/rstb.1998.0266
Pascual, R., Archer, M., Ortiz Jaureguizar, E., Prado, J. L., Godthelp, H., & Hand, S. J. (1992). First discovery of monotremes in South America. Nature, 356(6371), 704–706. https://doi.org/10.1038/356704a0
Pian, R., Archer, M., Hand, S. J., Beck, R. M. D., & Cody, A. (2016). The upper dentition and relationships of the enigmatic Australian Cretaceous mammal Kollikodon ritchiei. Memoirs of Museum Victoria, 74, 97–105. https://doi.org/10.24199/j.mmv.2016.74.10
Proske, U., & Gregory, J. E. (2003). Electrolocation in the platypus—Some speculations. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 136(4), 821–825. https://doi.org/10.1016/S1095-6433(03)00160-0
Rich, T. H., Vickers-Rich, P., Trusler, P., Flannery, T. F., Cifelli, R., Constantine, A., Kool, L., & van Klaveren, N. (2001). Monotreme nature of the Australian early Cretaceous mammal Teinolophos. Acta Palaeontologica Polonica, 46(1), 113–118.
Warren, W. C., Hillier, L. W., Marshall Graves, J. A., Birney, E., Ponting, C. P., Grützner, F., Belov, K., Miller, W., Clarke, L., Chinwalla, A. T., Yang, S.-P., Heger, A., Locke, D. P., Miethke, P., Waters, P. D., Veyrunes, F., Fulton, L., Fulton, B., Graves, T., … Wilson, R. K. (2008). Genome analysis of the platypus reveals unique signatures of evolution. Nature, 453(7192), 175–183. https://doi.org/10.1038/nature06936
Zhou, Y., Shearwin-Whyatt, L., Li, J., Song, Z., Hayakawa, T., Stevens, D., Fenelon, J. C., Peel, E., Cheng, Y., Pajpach, F., Bradley, N., Suzuki, H., Nand, A., Grützner, F., & et al. (2021). Platypus and echidna genomes reveal mammalian biology and evolution. Nature, 592(7856), 756–762. https://doi.org/10.1038/s41586-020-03039-0